Что значит "РНК вируса гепатита С не обнаружено"? Анализ РНК вируса гепатита С. Рнк-вирусы как причина развития рака - вирусный канцерогенез Есть ли у вирусов днк
ДНК-содержащие вирусы несут в качестве генетического материала либо одно -, либо двухцепочечную ДНК, которая может быть как линейной, так и кольцевой. В ДНК закодирована информация о всех белках вируса. Вирусы, заражающие бактерии, называются бактериофагами. К ДНК-содержащим вирусам относятся вирусы гепатита В, герпес, вирусы оспы, паповавирусы, гепаднавирусы, парвовирусы.
По виду цепи ДНК вирусы делятся на 3 группы:
Первая группа - вирусы с двуцепочечной ДНК. Репликация ДНК-генома этих вирусов осуществляется при посредстве промежуточных молекул РНК: Молекулы РНК образуются в результате транскрипции вирусных ДНК в клеточном ядре хозяйским ферментом ДНК-зависимой РНК-полимеразой. Транскрибируется только одна из нитей вирусной ДНК. Синтез ДНК на мРНК происходит в результате реакции, катализируемой обратной транскриптазой: сначала синтезируется (-) нить ДНК, а затем на вновь синтезированной (-) нити ДНК тот же фермент строит (+) нить.
Вторая группа - вирусы с двуцепочечной ДНК.В одних случаях производством как мРНК, так и ДНК занимаются клеточные ферменты; в других случаях вирусы используют собственные ферменты. Бывает, что те и другие ферменты обслуживают процесс репликации и транскрипции. К этой группе относятся вирусы герпеса, оспы и др.
Третья группа - вирусы с одноцепочечной ДНК, с негативной, либо с позитивной полярностью. Попав в клетку, вирусный геном сначала превращается в двуцепочечную форму, это превращение обеспечивает клеточная ДНК-зависимая ДНК-полимераза. Транскрипция и репликация на последующих этапах происходит так же, как и для вирусов, с (±) ДНК-геномом. Структура вируса: это молекула ДНК в белковой оболочке, называемой капсидом. Однако есть много разных вариантов строения вирусов: от просто покрытой белком ДНК до сложных макромолекулярных комплексов, окруженных мембранными структурами, например, вирус оспы. Если у вируса есть мембрана‚ говорят, что он в оболочке, а если мембраны нет, то вирус называют «раздетым». Различают четыре основных вида капсидов: спиральные, икосаэдрические, сложные без оболочки, сложные с оболочкой. Неизменным итогом заражения клеток ДНК-содержащими бактериофагами является лизис. ДНК-содержащие вирусы животных вызывают лизис редко, однако клетки могут погибнуть из-за возникших при заражении хромосомных повреждений, вследствие иммунологической реакции организма или просто в результате нарушения вирусом нормальных клеточных функций.
ДНК-содержащие опухолеродные вирусы разделяются на 5 классов:
* Полиомавирусы
– обезьяний вирус SV40, вирус полиомы мышей и вирусы человека ВК и JC.
* Папилломавирусы
– 16 вирусов папилломы человека и множество папилломовирусов животных.
* Аденовирусы
– 37 вирусов человека, множествоаденовирусов животных (например, 24 вируса обезьян и 9 вирусов крупного рогатого скота).
* Герповирусы
– вирусы простого герпеса человека, цитомегаловирус человека, вирус Эпштейна– Барр и онкогенные вирусы приматов, лошадей, кур, кроликов, лягушек.
* Вирусы, подобные вирусу гепатита В
, – вирус гепатита В человека, гепатита североамериканского сурка, гепатита земляных белок и гепатита уток.
IV. РНК-содержащие вирусы
РНК-содержащие микроорганизмы представлены гриппом и парагриппом, вирусом иммунодефицита человека (ВИЧ), гепатитом А парамиксовирусами, вирусами гриппа, коронавирусами, аренавирусами, ретровирусами, реовирусами, пикорнавирусами, капицивирусами, рабдовирусами, тогавирусами, флавивирусами и буньявирусами. РНК-содержащие вирусы не имеют ДНК, генетическая информация закодирована в РНК. Геномы почти всех известных РНК-содержащих вирусов - это линейные молекулы.
Геномы РНК-содержащих вирусов можно разделить на 3 группы.
Первая группа - это однонитевые геномы положительной полярности. Такие геномы обозначают как (+)РНК. Вирусные (+)РНК-геномы кодируют несколько белков. С помощью этого фермента синтезируются сначала (-) нити РНК фага, затем при наличии особого белка, называемого «хозяйским фактором», репликаза осуществляет синтез (+) нити РНК. На заключительной стадии из накопившихся вирусных белков и (+) РНК формируются вирионы. Упрощенная схема этого процесса такова:(+) РНК (-) РНК Инфекционный процесс состоит в проникновении вируса в растительную клетку с последующей быстрой утратой им капсида. Затем в результате трансляции непосредственно (+)РНК рибосомами клетки-хозяина образуются несколько белков, часть которых необходима для репликации вирусного генома.
Репликация осуществляется РНК-репликазой, продуцирующей копии РНК для новых вирионов. Синтез белка капсида происходит после того как инфицировавшая клетку РНК подвергается некоторой модификации, делающей возможным присоединение рибосом клетки к тому участку РНК, которым кодируется этот белок. Сборка вириона начинается с образования дисков из белка капсида. Два таких белковых диска образуют структуру, которая после связывания с ней РНК приобретает форму спирали. Присоединение молекул белка продолжается до тех пор, пока РНК не будет покрыта полностью. В окончательной форме вирион представляет собой цилиндр длиной 300 нм.
Вторая группа - это однонитевые геномы с негативной полярностью, т.е. (-)РНК-геномы.Поскольку (-)РНК не может выполнять функции мРНК, для образования «своих» мРНК вирус внедряет в клетку не только геном, но и фермент, умеющий снимать с этого генома комплементарные копии по схеме: (-) РНК (+) РНК. Этот вирусный фермент упакован в вирионе в удобной для доставки в клетку форме. Инфекционный процесс начинается с того, что вирусный фермент копирует вирусный геном, образуя (+) РНК, выступающую в качестве матрицы для синтеза вирусных белков, в том числе РНК-зависимой РНК-полимеразы, которая входит в состав образующихся вирионов. К вирусам с негативным РНК-геномом относятся: вирусы гриппа, кори, бешенства, желтой карликовости картофеля и др.
Третью группу составляют двунитевые геномы, (±) РНК-геномы. Известные двунитевые геномы всегда сегментированы, т.е. состоят из нескольких разных молекул, Сюда относятся реовирусы. Их размножение проходит по варианту, близкому к предыдущему. Вместе с вирусной РНК в клетку попадает и вирусная РНК-зависимая РНК-полимераза, которая обеспечивает синтез молекул (+) РНК. В свою очередь (+) РНК обеспечивает производство вирусных белков на рибосомах хозяйской клетки и служит матрицей для синтеза новых (-) РНК-цепочек вирусной РНК-полимеразой. Цепочки (+) и (-) РНК, комплексируясь друг с другом, образуют двунитевой (±) РНК-геном, который упаковывается в белковую оболочку. Реовирусы поражают респираторные и кишечные пути теплокровных животных (человека, обезьян, крупного и мелкого рогатого скота, летучих мышей.
Инфекционный процесс начинается с проникновения в клетку РНК. После частичного разрушения наружнего капсида ферментами лизосом РНК в образовавшейся таким образом субвирусной частице транскрибируется, ее копии покидают частицу и соединяются с рибосомами. Затем в клетке-хозяине продуцируются белки, необходимые для формирования новых вирусных частиц. Репликация РНК вирусов происходит по консервативному механизму. Одна из цепей каждого сегмента РНК служит матрицей для синтеза большого числа новых (+) цепей. На этих (+) цепях образуются затем как на матрице (–) цепи, (+) и (–) цепи при этом не расходятся, а остаются вместе в виде двухцепочечных молекул.
К РНК-содержащим вирусы также относятся вирусы, у которых цикл репликации генома можно разбить на две главные реакции: синтез РНК на матрице ДНК и синтез ДНК на матрице РНК. При этом в состав вирусной частицы в качестве генома может входить либо РНК, либо ДНК. Вирусная частица содержит две молекулы геномной одноцепочечной (+)РНК. В вирусном геноме закодирован необычный фермент, который обладает свойствами как РНК-зависимой, так и ДНК-зависимой ДНК-полимеразы.
V. Вирусные заболевания
Репродукция вирусов в природе поддерживается разными типами организмов: бактериями, грибами, простейшими, растениями, животными. Например, насекомые часто страдают от вирусов, которые накапливаются в их клетках в виде крупных кристаллов. Растения нередко поражаются мелкими и просто устроенными РНК-содержащими вирусами. Эти вирусы даже не имеют специальных механизмов для проникновения в клетку. Они переносятся насекомыми (которые питаются клеточным соком), круглыми червями и контактным способом, заражая растение при его механическом повреждении. Вирусы бактерий (бактериофаги) имеют наиболее сложный механизм доставки своего генетического материала в чувствительную бактериальную клетку. Сначала «хвост» фага, имеющий вид тонкой трубочки, прикрепляется к стенке бактерии. Затем специальные ферменты «хвоста» растворяют участок бактериальной стенки и в образовавшееся отверстие через «хвост», как через иглу шприца, впрыскивается генетический материал фага (обычно ДНК).
Более десяти основных групп вирусов патогенны для человека. Среди ДНК-содержащих вирусов это семейство поксвирусов (вызывающих натуральную оспу, коровью оспу и другие оспенные инфекции), вирусы группы герпеса (герпетические высыпания на губах, ветряная оспа), аденовирусы (заболевания дыхательных путей и глаз), семейство паповавирусов (бородавки и другие разрастания кожи), гепаднавирусы (вирус гепатита B). РНК-содержащих вирусов, болезнетворных для человека, значительно больше. Пикорнавирусы (от лат. pico – очень мелкий, англ. RNA – РНК) – самые мелкие вирусы млекопитающих, похожие на некоторые вирусы растений; они вызывают полиомиелит, гепатит А, острые простудные заболевания. Миксовирусы и парамиксовирусы – причина разных форм гриппа, кори и эпидемического паротита (свинки). Арбовирусы (от англ. arthropod borne – «переносимые членистоногими») – самая большая группа вирусов (более 300) – переносятся насекомыми и являются возбудителями клещевого и японского энцефалитов, желтой лихорадки, менингоэнцефалитов лошадей, колорадской клещевой лихорадки, шотландского энцефалита овец и других опасных болезней. Реовирусы – довольно редкие возбудители респираторных и кишечных заболеваний человека – стали предметом особого научного интереса в силу того, что их генетический материал представлен двухцепочечной фрагментированной РНК. Венерические болезни, ветряная оспа, гепатит, грипп, денге лихорадка, инфекционный мононуклеоз, корь, краснуха, менингит, оспа натуральная, полиомиелит, респираторные вирусные заболевания, свинка, Синдром приобретенного иммунодефицита (СПИД), энцефалит.
Возбудители некоторых болезней, в том числе очень тяжелых, не укладываются ни в одну из вышеперечисленных категорий. К особой группе медленных вирусных инфекций еще недавно относили, например, болезнь Крейтцфельда – Якоба и куру – дегенеративные заболевания головного мозга, имеющие очень продолжительный инкубационный период. Однако оказалось, что они вызываются не вирусами, а мельчайшими инфекционными агентами белковой природы – прионами.
Похожая информация.
Самые простые вирусы имеют в составе нуклеиновую кислоту, которая выступает в качестве генетического материала как самого микроорганизма, так и его капсида, представляющего собой чехол из белка. Состав некоторых вирусов дополнен жирами и углеводами. У вирусов отсутствует часть ферментов, которая отвечает за репродуктивную функцию, поэтому размножаться они могут, только попав в клетку живого организма. Метаболизм зараженной клетки после этого перестраивается на производство вирусных, а не собственных компонентов. В каждую клетку заложена определенная генетическая информация, которая при определенных обстоятельствах может рассматриваться в качестве инструкции по синтезу конкретного вида белка внутри клетки. Зараженная клетка воспринимает данную информацию как руководство к действию.

Размеры
Что касается размеров ДНК- и РНК-содержащих вирусов, то он находится в пределах 20-300 нм. Вирусы в большинстве своем имеют меньший размер, чем бактерии. Эритроцитные клетки, например, на порядок больше вирусных. Способная к заражению, полноценная инфекционная вирусная частица вне здорового организма носит название вирион. В ядро вириона входят одна или несколько молекул нуклеиновой кислоты. Капсид представляет собой оболочку из белка, которая покрывает вирионовую нуклеиновую кислоту, обеспечивая защиту от пагубного воздействия окружающей среды. Нуклеиновая кислота, входящая в вирион, считается геномом вируса и выражается в дезоксирибонуклеиновой кислоте, или ДНК, а также рибонуклеиновой (РНК). В отличие от бактерий, у вирусов не встречается комбинация этих двух видов кислоты.
Рассмотрим основные стадии размножения ДНК-содержащих вирусов.
Размножение вирусов
Чтобы иметь возможность необходимо внедриться в клетки носителя. Некоторые вирусы могут существовать у большого количества хозяев, у других имеется склонность к конкретным видам. На первоначальной стадии заражения вирус внедряет в клетку генетический материал в виде ДНК или РНК. Репродуктивная его функция, а также дальнейшее развитие клеток напрямую зависят от деятельности и выработки генов и белков вируса.
Для производства клеток ДНК-содержащим вирусам бывает недостаточно собственных белков, поэтому используются аналогичные вещества носителя. Спустя некоторое время после заражения в клетке остается только небольшая часть изначальных вирусов. Данная фаза называется эклипсной. Геном вируса в этот период тесно взаимодействует с носителем. Затем, спустя несколько этапов, начинается накопление во внутриклеточном пространстве потомства вируса. Это называется фазой созревания. Рассмотрим последовательность стадий размножения ДНК-содержащих вирусов.
Цикл жизни
Цикл жизни вирусов состоит из нескольких этапов, являющихся обязательными:
1. Адсорбция на клетке носителя. Это первоначальный и важный этап распознания клеток-мишеней по рецепторам. Адсорбция может происходить на клетках органов или тканей. Процесс запускает механизм дальнейшей интеграции вируса в клетку. Для связывания клеток требуется определенное количество ионов. Это необходимо для снижения электростатического отталкивания. Если проникновение в клетку не удается, вирус ищет новую мишень для интеграции и процесс повторяется. Это явление объясняет определенность в путях проникновения вируса в организм человека.
Так, например, слизистая оболочка верхних дыхательных путей обладает рецепторами к вирусу гриппа. Клетки кожи, напротив, таковых не имеют. По этой причине невозможно заразиться гриппом через кожный покров, это возможно только при вдыхании частиц вируса. Бактериальные вирусы в форме нитей или не имеющие отростков не могут прикрепляться на стенках клетки, поэтому они адсорбируются на фимфибриях. На начальном этапе адсорбция происходит за счет электростатического взаимодействия. Данная фаза обратима, так как частица вируса легко отделяется от клетки, выбранной в качестве мишени. Начиная со второй фазы, отделение не представляется возможным.

2. Следующая стадия размножения ДНК-содержащих вирусов характеризуется попаданием целого вириона или нуклеиновой кислоты, им выделяемой внутри клетки носителя . В организм животного вирус интегрируется проще, так как клетки в данном случае не снабжены оболочкой. Если у вириона есть снаружи липопротеидная мембрана, то она сталкивается при контакте с аналогичной защитой клетки носителя и вирус попадает в цитоплазму. Вирусы, проникающие в бактерии, растения и грибы интегрируются сложнее, так как в данном случае они вынуждены проходить сквозь жесткую стенку клетки. Для этого бактериофаги, к примеру, обеспечены ферментом лизоцимом, который помогает растворить твердые клеточные стенки. Примеры ДНК-содержащих вирусов рассмотрим ниже.
3. Третья стадия именуется депротеинизацией. Она характеризуется высвобождением нуклеиновой кислоты, являющейся носителем генетической информации. У некоторых вирусов, например, бактериофагов, данный процесс сочетается со второй стадией, так как белковая оболочка вириона остается за пределами клетки носителя. Вирион способен проникать в клетку путем захвата последней. При этом возникает вакуоль-фагосома, которая вбирает в себя первичные лизосомы. При этом расщепление на ферменты происходит только у белковой части вирусной клетки, а нуклеиновая кислота остается в неизменном состоянии. Именно она в дальнейшем существенно переформирует функционирование здоровой клетки, вынуждая ее производить нужные вирусу вещества. Сам вирус необходимыми для таких процедур механизмами не обеспечен. Существует такое понятие, как стратегия вирусного генома, которая подразумевает реализацию генетической информации.
4. Четвертая стадия размножения ДНК-содержащих вирусов сопровождается выработкой нужных для жизнедеятельности вируса веществ, которая осуществляется под влиянием нуклеиновой кислоты . Сначала вырабатывается ранняя мРНК, которая станет основой для белков вируса. Ранними называются молекулы, которые возникли до высвобождения нуклеиновой кислоты. Молекулы, возникшие после репликации кислоты, называют поздними. Важно понимать, что выработка молекул напрямую зависит от вида нуклеиновой кислоты конкретного вируса. ДНК-содержащие вирусы во время биосинтеза придерживаются определенной схемы, включающей конкретные этапы - ДНК-РНК-белок. Мелкие вирусы используют в процессе транскрипции РНК-полимеразы. Крупные, такие как оспяной вирус, синтезируются не в клеточном ядре, а в цитоплазме.
Группы РНК-вирусов
1. Первая группа устроена наиболее просто. В нее входят корона-, тога- и пикорнавирусы. Транскрипция у этих видов вируса не осуществляется, поскольку однонитчатая РНК вириона самостоятельно реализует функцию матричной кислоты, то есть представляет собой основу для выработки белков на уровне клеточных рибосом. Таким образом, схема биовыработки у них выглядит как РНК-белок. Вирусы данной группы называют также позитивно геномными или плюснитевыми.

2. Вторая группа ДНК и РНК-содержащих вирусов включает в себя минуснитевые вирусы, то есть они обладают негативным геномом. Это вирусы кори, гриппа, паротита и многие другие. В них также содержится однонитчатая РНК, но она не подходит для непосредственной трансляции. По этой причине сначала происходит перенос данных на РНК вириона, а полученная матричная кислота будет служить в дальнейшем основой для выработки белков вируса. Транскрипция в данном случае определяется зависимой от рибонуклеиновой кислоты полимеразой РНК. Данный фермент приносится вирионом, так как он отсутствует в клетке изначально. Это связано с тем, что у клетки не возникает необходимости перерабатывать РНК для получения другого РНК. Так, схема биовыработки в данном случае будет выглядеть как РНК-РНК-белок.
3. Третью группу составляют так называемые ретровирусы. Они же входят в разряд онковирусов. Биосинтез у них происходит по более сложному пути. В исходной однонитчатого типа на начальном этапе происходит выработка ДНК, что является уникальным явлением, аналогов которому нет в природе. Процесс находится под контролем специального фермента, а именно зависимой от РНК полимеразой ДНК. Данный фермент также носит название ревертазы или обратной транкриптазы. Молекула ДНК, полученная в результате биосинтеза, получает форму кольца и обозначается как провирус. Далее молекула внедряется в клетки хромосом носителя и транскрибируется несколько раз посредством полимеразы РНК. Созданные копии совершают следующие действия: представляют собой матрицу РНК, с помощью которой вырабатывается вирусный белок, а также вирион РНК. Схема синтеза представляется таким образом: РНК-ДНК-РНК-белок.
4. Четвертая группа образуется из вирусов, РНК которых имеет двухнитчатую форму. Их транскрипция реализуется посредством фермента вируса зависимой РНК полимеразы РНК.
5. В пятой группе выработка составляющих частицы вируса, а именно капсидных белков и нуклеиновой кислоты происходит многократно.
6. Шестая группа включает в себя вирионы, которые возникают как результат самосборки на основе множества копий белков и кислоты. С этой целью концентрация вирионов должна достичь критического значения. При этом компоненты частицы вируса вырабатываются отдельно друг от друга в разных областях клетки. Сложные вирусы также создают защитную оболочку из веществ, входящих в плазменную клеточную мембрану.
7. На заключительном этапе новые частицы вируса высвобождаются из клетки носителя. Данный процесс происходит разными способами в зависимости от вида вируса. Некоторые клетки после этого погибают, так как клеточный лизис освобождается. В других вариантах возможно отпочковывание от клетки, однако и этот способ не предотвращает ее дальнейшее отмирание, так как повреждается мембрана плазмы.

Период до момента выхода вируса из клетки называют латентным. Продолжительность данного промежутка может находиться в пределах от нескольких часов до пары дней.
Геномные вирусы, содержащие ДНК
1. Такие геномы, как адено-, папова- и герпесвирусов, переносятся и копируются в клеточном ядре носителя. содержащие двухцепочечную ДНК. Капсиды, попав в клетку, переносятся к мембране клеточного ядра, чтобы потом, под воздействием определенных факторов, ДНК вируса перешло в нуклеоплазму и накопилось там. Вирусы при этом пользуются матрицей РНК и клеточными ферментами носителя. А-белки переносятся первыми, за ними следуют b-белки и g-белки. Матрица РНК возникает на основе а-22 и а-47. реализует перенос ДНК, который размножается по принципу перекатывающегося кольца. Капсид, в свою очередь, возникает из g-5 белка. Какие еще существуют геномы ДНК-содержащих вирусов?
2. Поксивирусы входят во вторую группу. На начальном этапе действия осуществляются в цитоплазме. Там происходит высвобождение нуклеотидов и начало транскрипции. Затем образуется матрица РНК. На ранних стадиях выработки создается полимераза ДНК и около 70 белков, а также двухцепочная ДНК расщепляется полимеразой. С обоих сторон генома начинается репликация в тех местах, где на начальном этапе осуществлялось расплетение и расщепление цепей ДНК.
3. В третью группу включены парвовирусы. Размножение осуществляется в клеточном ядре носителя и находится в зависимости от функций клетки. В данном случае ДНК образует так называемую шпилечную структуру и выступает в роли затравки. 125 первых пар оснований переходят с начальной цепи на смежную, которая и служит матрицей. Таким образом, происходит инверсия. Для синтезирования нужна полимераза ДНК, благодаря которой происходит транскрипция вирусного генома.
8. В четвертую группу входят гепаднавирусы. Сюда относится ДНК-содержащий вирус гепатита. ДНК вируса кольцевого типа работает в качестве основы для выработки мРНК вируса и плюс-цепи РНК. Она, в свою очередь, становится матрицей для синтеза минус-цепи ДНК.
Методы борьбы

Как правило, антитела, направленные на борьбу с теми или иными вирусами вырабатываются как результат вторжения вредоносных микроорганизмов в систему носителя. Однако усилить выработку антител можно заранее, сделав профилактическую прививку.
Виды вакцинации
Существует несколько основных видов вакцинации, включающих:
1. Введение в организм ослабленных клеток вируса. Это провоцирует выработку увеличенного количества антител, что позволяет бороться с нормальным вирусным штаммом.
2. Введение уже мертвого вируса. Принцип действия аналогичен первому варианту.
3. Пассивная иммунизация. Данный метод заключается в введении уже синтезированных антител. Это может быть как кровь человека, перенесшего заболевание, против которого делается прививка, так и животного, например, лошадей. Последовательность размножения ДНК-содержащих вирусов мы рассмотрели.
Чтобы избежать заражения организма различными видами вирусов, опасных для здоровья человека, следует беречь тело от потенциального контакта с патогенными микроорганизмами. Вполне по силам избежать токсоплазмы, микоплазмы, герпеса, хламидий и других распространенных форм вируса, просто соблюдая определенные рекомендации. Это особенно касается детей до 15 лет.
Если тело ребенка не было заражено вышеперечисленными штаммами вирусов, то у него вырабатывается в подростковом периоде здоровый и усиленный иммунитет. Главная опасность вирусов - не всегда в том, как они выражаются, а в том, какое влияние оказывают на защитные свойства нашего организма. Примеры ДНК- и РНК-содержащих вирусов интересуют многих.

Герпесный вирус, который присутствует в теле 9 из 10 жителей Земли, снижает иммунные свойства примерно на 10 процентов в течение всей жизни, хотя может и никак не проявлять себя.
Заключение
Помимо подобной которая подчас не ограничивается только герпесом, условия современной жизни далеки от идеала, что также сказывается на защитных барьерах организма. В данный пункт можно отнести форсированный городской ритм жизни, плохую экологию, неправильное питание и т.д. На фоне снижения общего состояния здоровья человека его организм становится менее резистентным к различным вирусам и, соответственно, частым болезням.
Открытие вирусов , вызывающих злокачественные опухоли у животных, произошло еще на рубеже XIX и XX веков. В 1910 г. Пейтон Рауш обнаружил, что бесклеточный фильтрат из тканей птичьей саркомы может вызывать развитие аналогичной саркомы у цыплят. Примерно в это же время была доказана вирусная природа птичьего миелобластоза. Позднее было обнаружено, что часто проходит весьма значительный латентный период между инфицированием вирусом и развитием рака.
Тем не менее вплоть до 1960-х годов не существовало четких доказательств того, что инкорпорация вирусной ДНК в геном клетки является необходимым условием развития злокачественной трансформации, как не было и случаев выделения вирусной ДНК из раковых клеток.
В настоящее время выделяют два типа вирусных онкогенов . Оба этих типа онкогенов встраиваются в клеточную ДНК. Вирусы первого типа несут онкогены, которые вызывают быструю злокачественную «трансформацию» клеток в культурах in vitro, а в организме вызывают развитие опухолей. При втором типе вирус действует медленнее и для развития опухоли требуется значительное время. Вирусы второго типа не вызывают злокачественной трансформации клеток в культурах in vitro.
РНК-содержащие вирусы вызывают развитие целого ряда различных опухолей у животных, при этом наиболее распространена индукция этими вирусами лимфом, лейкемий и сарком. Типичная структура таких вирусов - это две идентичные цепочки молекул РНК в сочетании с ферментом обратная транскриптаза, одетых в гликопротеиновую оболочку. При инфицировании вирусом его обратная транскриптаза заставляет клетки синтезировать ДНК, комплементарную вирусной РНК.
Эта ДНК затем встраивается в клеточные хромосомы, и на ее основе клетка уже сама начинает синтез новых вирусных белков, вирусных обратных транскриптаз и элементов гликопротеиновой оболочки. Из-за механизма их действия этот тип вирусов получил название ретровирусы. Все они имеют весьма схожий внешний вид на электронных микрофотографиях и являются самыми маленькими из известных вирусов.
Некоторые из ретровирусов (например, вирусы птичьего лейкоза, кошачьей и мышиной лейкемии) содержат в себе только три гена и обладают очень длительным инкубационным периодом с момента заражения до возникновения опухоли. Другие вирусы (например, вирус саркомы Рауса (ВСР)) вызывают очень быструю злокачественную трансформацию и могут быть выделены из культуры опухолевых клеток.
Показано, что вирус ВСР содержит особый ген (v-src), способный вызывать трансформацию фибробластов in vitro. Этот ген кодирует наработку протеинкиназы, которая фосфорилирует тирозин. К сожалению, действие этой протеинкиназы запускает целый каскад различных метаболических процессов, и очень сложно оценить, какой именно из них ведет к злокачественной трансформации.
В настоящее время известно, что и нормальные, и злокачественные клетки содержат в своем генотипе участки ДНК, сходные или идентичные ряду последовательностей онкогенных РНК-содержащих вирусов. Такие участки получили название клеточных прото-онкогенов (чтобы отличать их от вирусных онкогенов). Постулируется, что активация этих участков, возникающая в результате канцерогенного воздействия, запускает целую цепочку событий, ведущую в итоге к злокачественной трансформации клетки. Считается также, что ретровирусы инкорпорировали эти клеточные участки в свой геном в процессе эволюции.
В настоящее время мы лучше понимаем механизмы действия продуктов вирусной активации . Примером такого продукта является протеинкиназа, активируемая геном sre, а также целый ряд других вирус-индуцированных канцерогенов. Это и рецепторы к эпидермальному фактору роста, продуцируемые геном v-erb, и тромбоцитарный фактор роста (ТФР), кодируемый фрагментами гена v-sis, и целый ряд связывающихся с ядром клетки белков, продукцию которых вызывает вирус птичьей лейкемии.
Вирусные онкогенные молекулы в большинстве случаев структурно отличаются от своих аналогов клеточного происхождения; кроме того, в них отсутствуют интроны. Например, белок, кодируемый геном v-erb, гомологичен клеточному рецептору к эпидермальному фактору роста (ЭФР), но в нем отсутствует часть экстраклеточного домена, включая ЭФК-специфичный сайт. Так как в продуцируемой вирусом молекуле отсутствует плазматический участок, ответственный за автофосфорилирование, такой вирусный рецептор все время находится во «включенном» состоянии.
Как нормальные, так и раковые клетки содержат в себе участки последовательностей ДНК, гомологичные РНК онкогенных вирусов. Если происходит экспрессия или активация под действием канцерогенов этих клеточных онкогенов, это ведет к злокачественной трансформации клеток.
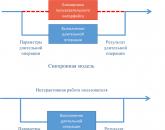
 Онкогенная и злокачественная трансформация.
Онкогенная и злокачественная трансформация.
На стадии А нормальная клетка, которая характеризуется низкой протоонкогенной активностью, продуцирует фактор роста (х) или белки дифференцировки или рецепторы (у).
Канцерогены повышают активность протоонкогенов, что дает начало неопластической трансформации.
По другому механизму: при заражении ретровирусом происходит внедрение в ДНК клетки вирусных промоторов или онкогенов (Б), что также ведет к увеличению онкогенной активности и последующей злокачественной трансформации.
Вирус может активировать процессинг в клетках путем внедрения в их специальных регуляторных последовательностей - промоторов считывания белка, нарушая таким образом нормальные процессы транскрипции. Запускаемый таким образом механизм внедренного мутагенеза может вовлекать в себя целый комплекс различных процессов. Примером может служить внедрение в ДНК клетки вирусной после довательности «многочисленных терминальных повторений» (МТП). При внедрении в клеточную ДНК данная последовательность инициирует транскрипцию в обоих направлениях цепочки ДНК, позволяя транскрибировать одновременно как клеточные, так и вирусные гены. Такой механизм действия характерен для вируса клеточного лейкоза, когда вирусная ДНК интегрируется в клеточную сразу вслед за участком с-тус, вызывая его активацию.
Первым ретровирусом , для которого была однозначно доказана связь со злокачественными новообразованиями, был вирус Т-клеточной лейкемии человека (ВТКЛ-1), выделенный из клеток хронической кожной Т-лимфомы. Этот вирус довольно широко распространен, может передаваться половым путем, через кровь, особенно в среде наркоманов, и от беременной женщины к плоду. Первоначально эндемический ареал циркуляции данного вируса был в основном представлен тропическими странами, однако в настоящее время в США серопозитивная реакция на вирус обнаруживается у каждого из 4000 человек населения. Кроме Т-клеточной лейкемии вирус вызывает тропический спастический паралич.
После 20 лет наблюдения за серопозитивными пациентами оценено, что риск развития последнего заболевания составляет около 5%. Один из вирусных генов, а именно tax-ген, вызывает увеличение продукции клеточного интерлейкина-2 (ИЛ-2) и его рецепторов, что является основным фактором, стимулирующим деление Т-клеток.
Ретровирусы могут вызывать опухолевые заболевания не напрямую, а опосредованно, как это было показано для вируса иммунодефицита человека (ВИЧ-1), вызывающего развитие СПИДа. Случаи развития рака у ВИЧ-инфицированных рассмотрены в одном из обзоров. Отмечается, что у ВИЧ-инфицированных наиболее часто развиваются три типа опухолей: мгновенная или высокозлокачественная В-клеточная лимфома; саркома Капоши (СК, которую вызывает другой вирус - герпесвирус ГСК, или герпесвирус 8); карцинома шейки матки.
До разработки эффективных методов терапии ВИЧ-инфицированных более 40% из них заболевали тем или иным видом рака. Тем не менее связь этого вируса с развитием онкологических заболеваний скорее всего косвенная и опосредована развитием общей хронической иммунодепрессии организма, которая и позволяет другим канцерогенным вирусам вызывать рак. В-клеточные лимфомы также характеризуются довольно сложным патогенезом. Хотя В-клетки не поражаются вирусом ВИЧ-1, они могут быть мишенью для других типов вирусов, например для вируса Эпштейна-Барра (ВЭБ). Рак шейки матки у женщин также развивается на основе вторичной вирусной инфекции - вируса папилломы человека (ВПЧ), на фоне общей иммунодепрессии, вызванной ВИЧ-1. По причине общего снижения иммунных реакций организма все эти опухоли развиваются особенно быстро и агрессивно.
Пикорна-, калици-, астро-, тога- и флавивирусы реплицируются наиболее коротким путем: их (+)РНК геном функционирует непосредственно как мРНК.
Геномы пикорна- и флавивирусов функционируют как единая полицистрон- ная мРНК, транслирующаяся прямо в единый полипротеин, который впоследствии расщепляется с образованием индивидуальных структурных и неструктурных белков. Одним из них является РНК-зависимая PH К-полимераза, которая реплицирует вирусный геном. Вирусная (+)РНК транскрибируется в комплиментарную (-)РНК-копию, которая служит матрицей для синтеза новых цепей (+)РНК (рис. 6), или может использоваться в качестве мРНК-матриц для синтеза новых минус-цепей и геномной РНК вирусного потомства.
Тога-, корона- и калицивирусы отличаются от пикорнавирусов тем, что на начальном этапе инфекции экспрессируется лишь часть геномной РНК с образованием белков. Последние осуществляют синтез минус-цепи, являющейся матрицей для синтеза различных по размеру классов молекул плюс-РНК. Полипротеины, образующиеся на коротких молекулах мРНК, расщепляются на структурные вирионные белки. Полноразмерные плюс-РНК упаковываются в вирионы.
У тогавирусов транслируется только около 2/3 вирусной РНК (5"-конец); образующийся полипротеин расщепляется на неструктурные белки, которые необходимы для транскрипции и репликации РНК. Вирусная РНК-полимераза синтезирует полноразмерную (-)РНК, на которой затем синтезируются два вида (+)РНК: полноразмерная вирионная РНК, предназначенная для включения в вирионы, и РНК, длина которой равна 1/3, и которая является колинеарной с 3"-концом вирусной РНК и транслируется в полипротеин, который расщепляется на структурные белки (рис. 7). У калицивирусов образуются полигеномные и субгеномные мРНК .
Корона- и артеривирусы демонстрируют необычную стратегию транскрипции: первоначально часть вирионной (+) РНК функционирует как мРНК и транслируется с образованием РНК-полимеразы, которая затем синтезирует полногеномную (-)РНК. На этой (-)РНК транскрибируется гнездо субгеном- ных мРНК с общими 3"-концами. Транслируются только 5"-концевые последовательности каждого члена этого гнезда транскриптов.
Главным отличием вирусов с позитивным геномом является их способность синтезировать ферменты, ответственные за репликацию вирусного генома. Поэтому РНК, выделенная из таких вирусов, инфекционна. Второе отличие состо-

ит в монолитности вирусного генома. Поэтому первичный продукт трансляции обеих РНК (геномной и мРНК) представляет собой единый белок, который в дальнейшем расщепляется на индивидуальные вирусные белки (в том числе структурные) .
Ортомиксо-, бунья- и аренавирусы. Геном представлен (-)РНК. Каждый геномный сегмент транскрибируется отдельно вирионной РНК-транскриптазой и транслируется в один или несколько белков. Эти вирусы характеризуются тем, что их геномная РНК выполняет две матричные функции: в процессе транскрипции и репликации. Транскрипция вирусного генома - первое событие после проникновения вируса в клетку, в результате которого образуются моноцис- тронные мРНК, кодирующие один белок. Репликацию начинают вновь синтезированные вирусные белки, приводящие к образованию плюс-цепи, которая служит матрицей для синтеза геномной (-)РНК (рис. 8).
Следовательно, плюс-транскрипт, функционирующий в качестве мРНК, отличается от (+)РНК, служащей матрицей для вирусного потомства, хотя и первый, и вторая синтезируются на геномной РНК.
Следует отметить, что S-сегмент РНК аренавирусов и некоторых буньявиру- сов является двуполярным, т.е. одна часть сегмента имеет (+) полярность, другая (-) полярность. Стратегия репликации двуполярных РНК-вирусов соответствует полярности их геномов и является смешанной, присущей стратегии репликации (+)РНК и (-)РНК вирусов .
Парамиксо-, рабдо-, фило- и борнавирусы имеют геномную одноцепочечную РНК и РНК-зависимую РНК-полимеразу (транскриптазу), которая транскрибирует пять или более субгеномных (+)РНК, каждая из которых служит в каче-

стве моноцистронной мРНК. В противоположность транскрипции в результате репликации (с помощью той же самой полимеразы, действующей в качестве репликазы) образуются полноразмерные (+)цепи РНК, которые служат матрицей для синтеза новых вирионных (-)цепей РНК. Стратегия репликации борна- вирусов в некоторых деталях является более сложной .
Рео- и бирнавирусы имеют сегментированный двуцепочечный РНК-геном. Негативные цепи каждого сегмента (2-12 сегментов) транскрибируются раздельно в цитоплазме вирионной транскриптазой с образованием мРН К. Эти позитивно полярные (+)РНК служат матрицей для репликации. Образовавшиеся двуспиральные РНК в свою очередь служат матрицей для дальнейшей транскрипции мРНК. Транскрипция генома реовирусов происходит внутри субвирусной частицы с помощью вирионной транскриптазы, причем копируется только одна нить каждого фрагмента двуцепочечной РНК (асимметричная транскрипция). Синтезирующиеся мРНК выходят из сердцевины вириона через 12 полых вершин частично разрушенного капсида. Таким образом, молекулы мРНК реовирусов выполняют две функции. Во-первых, они транслируются, обеспечивая синтез вирусных белков, и, во-вторых, они включаются (по одной молекуле мРНК каждого из 10--12 генов) в состав частиц-предшественниц вирионов. Каждая мРНК служит в этих частицах матрицей для синтеза комплементарной цепи, что ведет к образованию двуцепочечных сегментов вирусного генома (рис. 9).
Молекулы (+)РНК, синтезированные на родительской двуспиральной РНК, связываются с белками и образуют субвирусные частицы, в которых содержится 10-12 фрагментов (+)РНК. Фрагменты (+)РНК в составе субвирусных частиц служат матрицами для синтеза (-)РНК, которые в свободном виде в инфицированных клетках не обнаружены. Полагают, что транскрипция и репликация РНК

реовирусов осуществляется одним ферментом, который модифицируется в процессе репликации вируса .
Ретровирусы. Геном представлен двумя идентичными молекулами (+)РНК. Вместо того чтобы функционировать как мРНК, геном транскрибируется с помощью вирионной обратной транскриптазы, с образованием гибридной молекулы РНК-ДНК. Затем на (-)ДНК синтезируется (+)ДНК и образуется двуспиральная ДНК (другой активностью того же самого фермента) и непрерывно включается в клеточную ДНК. Интегрированная вирусная ДНК (провирус) транскрибируется клеточной РНК-полимеразой II с последующим сплайсингом РНК-транскриптов и расщеплением образующихся белков. Часть вновь синтезированных полноразмерных (+)РНК транскриптов соединяется в пары и образует диплоидные геномы новых вирионов (рис. 10).
Геном ретровирусов состоит из трех генов: (5" - 3") gag, pol и env, которые кодируют, соответственно, внутренние белки вируса, обратную транскриптазу и белки оболочки. Вирус саркомы Рауса имеет в своем геноме четвертый ген - онкоген (src), продукт которого играет ключевую роль в трансформации клеток, но не является необходимым для размножения вируса .
Организация генома митохондрий.
Существуют два типа цитоплазматических ДНК: одни находятся в митохондриях эукариот, другие -в хлоропластах зеленых растений и водорослей. Как и все цитоплазматические элементы, они наследуются по материнской линии, а не по законам Менделя! Большая часть белков этих органелл, закодированная в ядерной ДНК, синтезируется в цитоплазме и затем переходит в органеллу. Однако некоторые белки митохондрий и хлоропластов и все их РНК кодируется в ДНК самих органелл и в них же синтезируются. Таким образом, органеллы - это результат объединенных усилий двух геномов и двух трансляционных аппаратов. РНК-компоненты рибосом органелл, а также тРНК, использующиеся при трансляции, кодируются геномами митохондрий и хлоропластов.
Размеры генома хлоропластов у всех исследованных организмов сходны, тогда как митохондриальные геномы у растений намного больше, чем у животных.
Все митохондрии и хлоропласты содержат по несколько копий собственной геномной ДНК. Эти молекулы ДНК обычно распределены в виде отдельных групп в матриксе митохондрий и в строме хлоропластов, где они прикреплены к внутренней мембране. Способ упаковки ДНК неизвестен. По структуре геном более сходен с бактериальным геномом: например, как и у бактерий, у них нет гистонов.
Геном вирусов включает:
– Структурные гены, которые кодируют белки. Занимают примерно 95 % вирусной хромосомы. Белки вирусов можно разделить на несколько групп: структурные, ферменты, регуляторы.
– Регуляторные последовательности, которые не кодируют белки: промоторы, операторы и терминаторы.
– Прочие некодирующие участки (сайты), в том числе:
– участок attP, обеспечивающий интеграцию вирусной хромосомы в хромосому клетки–хозяина;
– участки cos – липкие концевые участки линейных вирусных хромосом, обеспечивающие замыкание линейной хромосомы в кольцевую форму.
Гены, кодирующие рРНК и тРНК, в геноме вирусов обычно отсутствуют. Однако в геноме крупного фага Т4 имеются гены, кодирующие несколько тРНК.
Геном вирусов отличается высокой плотности упаковки информации. Например, у фага φХ174 в пределах одного гена может располагаться еще один ген. В частности, ген В находится в пределах гена А, а ген Е – в пределах гена D. У мелкого РНК-содержащего фага f2 ген регуляторного белка, блокирующего лизис (созревание вирионов и разрушение клетки), перекрывается с двумя другими генами, удаленными друг от друга.
Особенности вирусов эукариот
У вирусов эукариот обнаружены следующие особенности:
1. Интрон-экзонная структура генов.
2. Модификация белков после синтеза полипротеинов: весь геном транскрибируется в виде одной молекулы мРНК, которая служит матрицей для синтеза полипротеина – одного гигантского инертного белка, и лишь затем происходит расщепление полипротеина на белки, выполняющие определенные функции.
3. Перекрывание генов (обезьяний вирус SV 40, вирус гриппа).
К ДНК-содержащим вирусам относятся многие вирусы бактерий – бактериофаги (или просто фаги). Некоторые мелкие фаги (например, фаг М13) при репродукции не разрушают клетку. Репродукция крупных фагов (например, фага Т–4) приводит к гибели клетки. Фаг Т–4 – это один из наиболее сложно организованных вирусов. Белковый капсид включает не менее 130 белков, образующих головку, воротничок, сократимый хвост, базальную пластинку и хвостовые нити. Такое строение капсида позволяет впрыскивать ДНК в бактериальную клетку через толстую оболочку, поэтому подобные вирусы образно называют «живыми шприцами». Т–фаги могут существовать в виде профага длительное время. К ДНК-содержащим вирусам относятся возбудители многих заболеваний человека и животных: вирусы оспы, герпеса, гепатита В, аденовирусы млекопитающих и человека (вызывают желудочно-кишечные заболевания, ОРВИ, конъюнктивиты), вирусы бородавок человека. К ДНК-содержащим вирусам относятся и некоторые вирусы растений (вирус золотистой мозаики фасоли, вирус мозаики цветной капусты). Некоторые вирусы используются в генной инженерии для переноса генов от одних организмов к другим, например, обезьяний вирус SV 40.
Вирионы ДНК-содержащих вирусов содержат ДНК. Объемом ДНК определяется количество белков в вирионе: один полипептид кодируется отрезком ДНК длиной примерно 1 тысяча нуклеотидов (нуклеотидных пар). После проникновения в клетку вирусная ДНК становится матрицей для синтеза ДНК и РНК.
Примеры организации генома ДНК-содержащих вирусов
1. Кольцевая двухцепочечная ДНК длиной около 5 тпн.
– Обезьяний вирус SV 40. Мелкий эукариотический вирус. Вирионы в виде икосаэдра. Капсид белковый. Используется в генной инженерии как вектор переноса генов. Кодирует 5 белков.
– Вирусы бородавок человека.
2. Кольцевая одноцепочечная ДНК длиной около 5 тн; может быть как кодирующей, так и антикодирующей.
– Мелкие бактериофаги типа М13. Не разрушают клетку. Капсид включает 8 белков.
– Вирус золотистой мозаики фасоли.
3. Линейная двухцепочечная ДНК длиной 30-150 тпн.
– Бактериофаги типа Т4. Вирионы крупные. Белковый капсид из 130 белков включает: головку, хвостовой отдел и хвостовые нити. Эти вирусы могут существовать в виде профага длительное время.
– Аденовирусы млекопитающих и человека. Вирионы средних размеров в виде икосаэдра. Капсиды белковые. Вызывают ОРВИ, конъюнктивиты, желудочно-кишечные заболевания, иногда обладают онкогенными свойствами.
– Вирусы оспы, герпеса и им подобные. Вирионы крупные. Имеется липопротеиновая оболочка.
4. Линейная одноцепочечная ДНК длиной около 5 тн; ДНК может быть как кодирующей, так и антикодирующей. У человека известны как спутники аденовирусов.
5. Двухцепочечная ДНК, замкнутая в кольцо из перекрывающихся сегментов. Длина ДНК – 3-8 тн.
– Вирус гепатита В. Вирион сферический, средних размеров. Имеется дополнительная оболочка из вирусных и клеточных белков. Кодирует 5 белков.
– Вирус мозаики цветной капусты (CaMV). Промотор 35S-RNA (CaMV35S) этого вируса широко используется в традиционной генной инженерии для создания генетических конструкций.
К РНК-содержащим вирусам относятся многие вирусы растений, возбудители заболеваний человека и животных: вирус полиомиелита, вирусы гриппа А, В и С, вирусы паротита (свинки), кори, чумы плотоядных животных (чумки), бешенства, вирус иммунодефицита человека (ВИЧ). В отдельную группу выделяются арбовирусы, которые переносятся членистоногими (клещами, москитами), например, вирусы клещевого энцефалита, желтой лихорадки. Многие РНК-содержащие вирусы вызывают ОРВИ (например, коронавирусы), желудочно-кишечные заболевания (реовирусы птиц, млекопитающих и человека). Некоторые РНК-содержащие вирусы используются в биотехнологии, например, вирусы полиэдроза насекомых.
Вирионы РНК-содержащих вирусов содержат РНК. После проникновения в клетку вирусная РНК становится матрицей для синтеза ДНК и РНК.
Примеры организации генома РНК-содержащих вирусов
1. Линейная одноцепочечная мРНК (плюс–цепь) длиной около 4 тн; в виде единой молекулы или в виде нескольких разных молекул. Плюс-цепь сразу же может использоваться для трансляции. Вегетативно-репродуктивная фаза этих вирусов протекает в цитоплазме. В плюс-цепи закодирована РНК-репликаза (РНК-зависимая РНК-полимераза). Представители:
– Вирус табачной мозаики (ВТМ) – сегментированная РНК. Вирион нитевидный (18х300 нм). ВТМ открыт Д.И. Ивановским в 1982 г.
– Вирус полиомиелита – несегментированная РНК. Вирионы мелкие, в виде икосаэдра. Капсид белковый.
– Вирус бешенства. Нитевидный вирион. Имеется дополнительная липопротеиновая оболочка.
– Арбовирусы (переносятся членистоногими: клещами, москитами) – вирусы клещевого энцефалита, желтой лихорадки. Морфология и размеры вирионов разнообразны, например, вирус энцефалита содержит 9 белков. Имеется дополнительная липопротеиновая оболочка.
– Мелкие бактериофаги (с несегментированной РНК).
2. Линейная одноцепочечная кРНК (минус–цепь, порядок нуклеотидов комплементарен по отношению к мРНК). Минус–цепь не может служить для трансляции и используется как матрица для синтеза плюс–цепи. Плюс-цепь служит для трансляции вирусных белков и используется как матрица для синтеза вирусной кРНК. Вегетативно-репродуктивная фаза этих вирусов также протекает в цитоплазме.
– Вирусы гриппа А, В, С. Вирус гриппа А содержит минус-цепь РНК, состоящую из 8 фрагментов. Фрагменты РНК связаны с вирусными белками и образуют спиральный нуклеокапсид. Поверх нуклеокапсида располагается гликолипопротеиновый суперкапсид. В составе вириона 10 белков. В состав суперкапсида входит два белка, определяющих антигенные свойства вируса: гемагглютинин и нейраминидаза. Кроме того, в состав вириона входит уже готовая РНК-репликаза, обеспечивающая синтез плюс-цепи на матрице минус-цепи.
– Вирусы паротита (свинки), кори, чумы плотоядных животных (чумки). Сферический вирион средних размеров. Имеется дополнительная липопротеиновая оболочка.
3. Линейная двухцепочечная РНК
– Мелкие бактериофаги. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый.
– Вирусы полиэдроза насекомых. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Используются в биотехнологии (для синтеза интерферона).
– Реовирусы птиц, млекопитающих и человека. Вирионы мелкие, сферические или в виде икосаэдра. Капсид белковый. Вызывают ОРВИ, желудочно-кишечные заболевания. РНК фрагментированная (10...11 фрагментов), кодирует 11 белков.
4. Две линейные одноцепочечные одинаковые молекулы мРНК длиной около 10 тн. Ретровирусы. Способны интегрироваться в ДНК. В состав вирионов входит фермент обратная транскриптаза (ревертаза). Имеется дополнительная липопротеиновая оболочка. Многие ретровирусы вызывают онкологические заболевания: лейкозы, саркомы, опухоли молочных желез. К ретровирусам относится и вирус иммунодефицита человека, вызывающий СПИД.
– Вирус иммунодефицита человека (ВИЧ). Содержит одну плюс-цепь РНК, кодирует 13 белков. Сферический вирион. Имеется дополнительная липопротеиновая оболочка, включающая фрагменты мембран человека. Избирательно поражает Т–лимфоциты.


Популярное
- Отсутствует файл d3dx9_43
- Тексты песен Lana Del Rey
- Как восстановить поврежденные документы Microsoft Word и файлы с ошибкой чтения Как открыть поврежденный файл docx
- Freebitcoin — регистрация, как заработать первый биткоин Фрее биткоин
- Получение root на планшете lenovo
- Explay Golf не включается, выключается, зависает?
- Лучшие безрамочные смартфоны
- Android Шаг за Шагом: ClockWork MOD Recovery - Что это и зачем он?
- Установка официальной прошивки на Samsung Galaxy A7 (2016)
- Как удалить проблемные обновления Windows Удалить обновления windows 7